Exam: Rozdiel medzi revíziami
(→Expresia génov, regulácia, motívy) |
(→Additional important concepts for computer scientists) |
||
| (26 intermediate revisions by the same user not shown) | |||
| Riadok 1: | Riadok 1: | ||
== Exam rules == | == Exam rules == | ||
| − | The | + | Exam is for 1-BIN-301 (mainly AIN, BIN, DAV, INF students). |
| + | |||
| + | The exam is primarily '''written''': | ||
* You need at least 50% of points | * You need at least 50% of points | ||
* Time 3 hours | * Time 3 hours | ||
| − | * About 50% of points for simple questions | + | * About 50% of points for simple questions |
** examples on this page | ** examples on this page | ||
** in case of interest tutorial session before exam | ** in case of interest tutorial session before exam | ||
* The rest of the questions mostly designing/modifying an algorithm or model | * The rest of the questions mostly designing/modifying an algorithm or model | ||
| − | |||
* You can use pen, simple calculator and a cheat sheet up to 2 A4 two-sided sheets | * You can use pen, simple calculator and a cheat sheet up to 2 A4 two-sided sheets | ||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
<!-- | <!-- | ||
| Riadok 41: | Riadok 16: | ||
--> | --> | ||
| − | + | If you do not pass the first written exam, part of your exam may be oral on the second and third attempts. | |
| − | + | ==Syllabus and examples of simple questions== | |
| − | + | Below we list the most important concepts that both biologists and computer scientists should know from this course. | |
| + | We also list simple questions. Questions of this type will comprise approximately 50% of the exam. Not all of these questions will be used on the exam and particular string, numbers, grammars and sequences will differ. | ||
===Sequencing and genome assembly=== | ===Sequencing and genome assembly=== | ||
| + | |||
| + | '''Main concepts in English and Slovak''' | ||
DNA sequencing and its use, sequencing read, paired reads, contigs, shortest common superstring problem, de Bruijn graphs | DNA sequencing and its use, sequencing read, paired reads, contigs, shortest common superstring problem, de Bruijn graphs | ||
Sekvenovanie DNA a jeho využitie, čítanie (read), spárované čítania, kontig, problém najkratšieho spoločného nadslova, de Bruijnove grafy | Sekvenovanie DNA a jeho využitie, čítanie (read), spárované čítania, kontig, problém najkratšieho spoločného nadslova, de Bruijnove grafy | ||
| + | |||
| + | '''Simple questions for the exam''' | ||
* Find the shortest common superstring of strings GACAATAA, ATAACAC, GTATA, TAATTGTA. | * Find the shortest common superstring of strings GACAATAA, ATAACAC, GTATA, TAATTGTA. | ||
| Riadok 59: | Riadok 39: | ||
===Sequence alignment=== | ===Sequence alignment=== | ||
| − | The problem of local and global alignment of two sequences, dynamic programming algorithms, scoring matrix and its probabilistic meaning, statistical significance (E-value, P-value), heuristic search of local alignments (BLAST), whole-genome and multiple alignments | + | '''Main concepts in English and Slovak''' |
| + | |||
| + | The problem of local and global alignment of two sequences, dynamic programming algorithms, scoring matrix and its probabilistic meaning, statistical significance (E-value, P-value), heuristic search of local alignments (BLAST, minimizers), whole-genome and multiple alignments | ||
| + | |||
| + | Problém lokálneho a globálneho zarovnania dvoch sekvencií, jeho riešenie pomocou dynamického programovania, skórovacia matica a jej pravdepodobnostný význam, štatistická významnosť (E-value, P-value), heuristické hľadanie lokálnych zarovnaní (BLAST, minimizer-y), celogenómové a viacnásobné zarovnania | ||
| − | + | '''Simple questions for the exam''' | |
| − | * Fill in the dynamic programing matrices for local and global alignment of sequences TACGT | + | * Fill in the dynamic programing matrices for local and global alignment of sequences TACGT and CAGGATT, where the match has score +3, mismatch -1, gap -2. Reconstruct also the optimal alignments found by the dynamic programming algorithm. |
* Compute the score of the alignment shown below using the scoring matrix shown below, gap opening penalty -5, gap extension penalty -2 for each additional base. Find a global alignment with a higher score for these two sequences and compute its score. (It is not necessary to find the optimal alignment; you can use any method to arrive at the answer.) | * Compute the score of the alignment shown below using the scoring matrix shown below, gap opening penalty -5, gap extension penalty -2 for each additional base. Find a global alignment with a higher score for these two sequences and compute its score. (It is not necessary to find the optimal alignment; you can use any method to arrive at the answer.) | ||
| Riadok 80: | Riadok 64: | ||
===Gene finding=== | ===Gene finding=== | ||
| − | + | '''Main concepts in English and Slovak''' | |
| − | + | Gene, exon, intron, mRNA, splicing and alternative splicing, genetic code, hidden Markov model (HMM), its states, transition and emission probabilities, use of HMMs in gene finding, Viterbi and forward algorithms | |
| + | Gén, exón, intrón, mRNA, zostrih a alternatívny zostrih, kodón, genetický kód, skrytý Markovov model (HMM), jeho stavy, pravdepodobnosti prechodu a emisie, použitie HMM na hľadanie génov, Viterbiho a dopredný algoritmus | ||
| − | * What is the | + | '''Simple questions for the exam''' |
| + | |||
| + | * What is the probability of generating sequence AGT using sequence of states 1,2,1 in the HMM below? | ||
<pre> | <pre> | ||
The HMM has three states 1, 2, 3. It always starts in state 1. | The HMM has three states 1, 2, 3. It always starts in state 1. | ||
| Riadok 106: | Riadok 93: | ||
===Evolution and comparative genomics=== | ===Evolution and comparative genomics=== | ||
| − | + | '''Main concepts in English and Slovak''' | |
| − | + | Phylogenetic tree (rooted and unrooted), maximum parsimony method, neighbor joining method, maximum likelihood method, Jukes-Cantor substitution model and more complex substitution matrices, Felsenstein algorithm, homolog, paralog, ortholog, detection of positive and negative selection, phylogenetic HMMs, likelihood ratio test | |
| − | * Find the most parsimonious assignment of bases at the ancestral nodes in the tree below given a column of alignment TTAAA (in the order gollum, hobbit, human, elf, orc). You can derive your answer using any method. | + | Fylogenetický strom (zakorenenený a nezakorenený), metóda maximálnej úspornosti (parsimony), metóda spájania susedov (neighbor joining), metóda maximálnej vierohodnosti (maximum likelihood), Jukes-Cantorov model substitúcií a zložitejšie substitučné matice, Felsensteinov algoritmus, homológ, paralóg, ortológ, detekcia pozitívneho a negatívneho výberu, fylogenetické HMM, test pomerom vierohodností (likelihood ratio test) |
| + | |||
| + | '''Simple questions for the exam''' | ||
| + | |||
| + | * Find the most parsimonious assignment of bases at the ancestral nodes in the tree below given a column of alignment TTAAA (in the order gollum, hobbit, human, elf, orc). You can derive your answer using any method (but it has to be the most parsimonious assignment). | ||
<pre> | <pre> | ||
Gollum ----| | Gollum ----| | ||
| Riadok 123: | Riadok 114: | ||
</pre> | </pre> | ||
| − | * Find the most parsimonious tree for the alignment given below. What is its cost (i.e. how many mutations are necessary to explain these sequences)? You can derive your answer using | + | * Find the most parsimonious tree for the alignment given below. What is its cost (i.e. how many mutations are necessary to explain these sequences)? You can derive your answer using any method (but it has to be the most parsimonious tree). |
| − | any method. | + | |
<pre> | <pre> | ||
whitebird ACAACGTCT | whitebird ACAACGTCT | ||
| Riadok 133: | Riadok 123: | ||
</pre> | </pre> | ||
| − | * Consider the tree for gollum, hobbit etc. given above, where each branch has the same length t. Let us assume that for any two different bases x and y, the probability of base x mutating to base y over time y is 0.1, and thus the probability of base x remaining the same after time t is 0.7. Probability of each base in the root is 0.25. Compute the probability that the tree will have base A in all internal nodes and in leaves bases TTAAA (from top to bottom). Find an | + | * Consider the tree for gollum, hobbit etc. given above, where each branch has the same length t. Let us assume that for any two different bases x and y, the probability of base x mutating to base y over time y is 0.1, and thus the probability of base x remaining the same after time t is 0.7. Probability of each base in the root is 0.25. Compute the probability that the tree will have base A in all internal nodes and in leaves bases TTAAA (from top to bottom). Find an assignment of bases in the ancestral nodes with a higher probability and compute this probability (you do not need to find the best possible assignment). |
| Riadok 145: | Riadok 135: | ||
</pre> | </pre> | ||
| − | === | + | ===Gene expression, regulation, motifs=== |
| − | + | '''Main concepts in English and Slovak''' | |
| − | Určovanie génovej expresie pomocou microarray alebo sekvenovaním RNA-seq, hierarchické zhlukovanie, klasifikácia, reprezentácia sekvenčných motívov (väzobné miesta transkripčných faktorov) ako konsenzus, regulárny výraz a PSSM, hľadanie nových motívov v sekvenciách, consensus pattern problem, hľadanie motívu pomocou pravdepodobnostných modelov | + | Measuring gene expressing using microarray or RNA-seq, hierarchical clustering, classification, representation of sequence motifs (transcription factor binding sites) as a consensus, regular expression and PSSM, finding new motifs in sequences, consensus pattern problem, finding motifs using probability models, EM algorithm |
| + | |||
| + | Určovanie génovej expresie pomocou microarray alebo sekvenovaním RNA-seq, hierarchické zhlukovanie, klasifikácia, reprezentácia sekvenčných motívov (väzobné miesta transkripčných faktorov) ako konsenzus, regulárny výraz a PSSM, hľadanie nových motívov v sekvenciách, consensus pattern problem, hľadanie motívu pomocou pravdepodobnostných modelov, algoritmus EM | ||
| + | '''Simple questions for the exam''' | ||
| + | |||
* After a series of expression measurements for 5 genes, we have computed distances between pairs of expression profiles and obtained the distance table shown below. Find the hierarchical clustering of these genes, where the distance between two clusters is computed as the minimum of the closest genes in these clusters (single linkage clustering). Show the order in which you were creating individual clusters. | * After a series of expression measurements for 5 genes, we have computed distances between pairs of expression profiles and obtained the distance table shown below. Find the hierarchical clustering of these genes, where the distance between two clusters is computed as the minimum of the closest genes in these clusters (single linkage clustering). Show the order in which you were creating individual clusters. | ||
<pre> | <pre> | ||
| Riadok 161: | Riadok 155: | ||
</pre> | </pre> | ||
| − | * Consider a motif represented by | + | * Consider a motif represented by the PSSM shown below. Compute the score of string GGAG. Which sequences of length 4 will have the smallest and highest score in this PSSM? |
<pre> | <pre> | ||
A -3 3 -2 -2 | A -3 3 -2 -2 | ||
| Riadok 168: | Riadok 162: | ||
T 1 -1 1 -2 | T 1 -1 1 -2 | ||
</pre> | </pre> | ||
| + | |||
| + | * We are running the EM algorithm for finding matrix W of base frequencies at individual motif positions. Motif length is 3 and we have 2 sequences of length 5 each, so there are 3 positions where the motif can start in each sequence. For each of these positions, we calculated the probability that the motif starts there based on the previous version of matrix W. What will be the new matrix W? (We do not use pseudocounts here, so for example base G will have probability 0 everywhere.) | ||
| + | |||
| + | <pre> | ||
| + | Sequence 1: AACAT, | ||
| + | probabilities of motif starting at positions 0,1,2: 0.4, 0.2, 0.4 | ||
| + | |||
| + | Sequence 2: AAAAT, | ||
| + | probabilities of motif starting at positions 0,1,2: 0.2, 0.2, 0.6 | ||
| + | </pre> | ||
| + | |||
* Find all occurrences of regular expression TA[CG][AT]AT in sequence GACGATATAGTATGTACAATATGC. | * Find all occurrences of regular expression TA[CG][AT]AT in sequence GACGATATAGTATGTACAATATGC. | ||
| − | === | + | ===Proteins=== |
| − | + | '''Main concepts in English and Slovak''' | |
| − | + | Primary, secondary and tertiary structure of a protein, protein domains and families, family representation by a profile (PSSM) and a profile HMM, protein threading, gene ontology. | |
| + | |||
| + | Primárna, sekundárna a terciálna štruktúra proteínov, proteínové domény a rodiny, reprezentovanie rodiny pravdepodobnostným profilom a profilovým HMM, protein threading, gene ontology. | ||
| + | |||
| + | '''Simple questions for the exam''' | ||
| + | |||
| + | * Construct a profile (PSSM) for the sequence alignment shown below, assuming that amino acid A comprises 60% of all sequences in a database, G 40% and we do not consider other amino acids. Use natural logarithm (ln) and pseudocount 1. | ||
<pre> | <pre> | ||
AAGA | AAGA | ||
| Riadok 186: | Riadok 197: | ||
===RNA=== | ===RNA=== | ||
| − | Sekundárna štruktúra RNA, pseudouzol a dobre uzátvorkovaná štruktúra, Nussinovovej algoritmus, minimalizácia energie, stochastické bezkontextové gramatiky, kovariančné modely. | + | '''Main concepts in English and Slovak''' |
| + | |||
| + | Secondary structure of RNA, pseudoknot and well-parenthesized structures, Nussinov algorithm, energy minimization, stochastic context-free grammars, covariance models of RNA families | ||
| + | |||
| + | Sekundárna štruktúra RNA, pseudouzol a dobre uzátvorkovaná štruktúra, Nussinovovej algoritmus, minimalizácia energie, stochastické bezkontextové gramatiky, kovariančné modely rodín RNA. | ||
| + | |||
| + | '''Simple questions for the exam''' | ||
| − | * | + | * Fill in the missing values in the matrix of dynamic programming for Nussinov algorithm which finds the RNA secondary structure without pseudoknots with the highest number of paired bases in RNA sequence GAACUAUCUGA (we allow only complementary bases A-U, C-G) and show the secondary structure found by the algorithm. |
<pre> | <pre> | ||
0 0 0 1 1 2 2 3 3 ? ? | 0 0 0 1 1 2 2 3 3 ? ? | ||
| Riadok 203: | Riadok 220: | ||
</pre> | </pre> | ||
| − | * | + | * Consider RNA sequence of length 27, which has secondary structure pairs on positions (2,23), (3,22), (4,21), (5,13), (6,12), (8,16), (9,15), (10,14), (18,26), (19,25). What is the smallest number of pairs that needs to be removed from this list to get a structure without pseudoknots? Which pairs will be removed? |
| − | * | + | * Consider RNA sequence ACUGAGUCCAAGG, which has secondary structure pairs on positions (1,7), (2,6), (3,5), (8,13) a (9,12). (Positions are numbered started from 1.) (This RNA is shown as an example in the lecture on RNA structure.) Show a derivation of this sequence using a grammar show below so that paired bases are always generated in the same step of the derivation. |
| − | ** | + | ** Grammar 1: S->aSu|uSa|cSg|gSc|aS|cS|gS|uS|Sa|Sc|Sg|Su|SS|epsilon |
| − | ** | + | ** Grammar 2: S->aSu|uSa|cSg|gSc|TS|ST|SS|epsilon; T->aT|cT|gT|tT|epsilon |
| − | === | + | ===Population genetics=== |
| + | |||
| + | '''Main concepts in English and Slovak''' | ||
| + | |||
| + | Polymorphism, SNP, allele, homozygote, heterozygote, recombination, allele frequency as a Markov chain, random genetic drift, linkage disequilibrium, association mapping, LD block, subpopulation. | ||
Polymorfizmus, SNP, alela, homozygot, heterozygot, rekombinácia, frekvencia polymorfizmu ako markovovský reťazec, náhodný genetický drift, väzbová nerovnováha (linkage disequilibrium), mapovanie asociácií, LD blok, subpopulácia. | Polymorfizmus, SNP, alela, homozygot, heterozygot, rekombinácia, frekvencia polymorfizmu ako markovovský reťazec, náhodný genetický drift, väzbová nerovnováha (linkage disequilibrium), mapovanie asociácií, LD blok, subpopulácia. | ||
| − | * | + | '''Simple questions for the exam''' |
| + | |||
| + | * For pairs of SNPs from the tables show below determine, if we can statistically reject the hypothesis that they are in linkage equilibrium (LE) at the significance level p=0.05, i.e. <math>\chi^2>3.841</math>. For each pair compute their <math>\chi^2</math> value. | ||
<pre> | <pre> | ||
Q q Q q Q q | Q q Q q Q q | ||
| Riadok 220: | Riadok 243: | ||
</pre> | </pre> | ||
| − | === | + | ===Additional important concepts=== |
| − | + | * Advanced dynamic programming algorithms (protein MS/MS, variants of sequence alignment, variants of Nussinov algorithm) | |
| − | * | + | * Integer linear programming |
| − | * | + | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
Aktuálna revízia z 15:27, 7. december 2023
Obsah
Exam rules
Exam is for 1-BIN-301 (mainly AIN, BIN, DAV, INF students).
The exam is primarily written:
- You need at least 50% of points
- Time 3 hours
- About 50% of points for simple questions
- examples on this page
- in case of interest tutorial session before exam
- The rest of the questions mostly designing/modifying an algorithm or model
- You can use pen, simple calculator and a cheat sheet up to 2 A4 two-sided sheets
If you do not pass the first written exam, part of your exam may be oral on the second and third attempts.
Syllabus and examples of simple questions
Below we list the most important concepts that both biologists and computer scientists should know from this course.
We also list simple questions. Questions of this type will comprise approximately 50% of the exam. Not all of these questions will be used on the exam and particular string, numbers, grammars and sequences will differ.
Sequencing and genome assembly
Main concepts in English and Slovak
DNA sequencing and its use, sequencing read, paired reads, contigs, shortest common superstring problem, de Bruijn graphs
Sekvenovanie DNA a jeho využitie, čítanie (read), spárované čítania, kontig, problém najkratšieho spoločného nadslova, de Bruijnove grafy
Simple questions for the exam
- Find the shortest common superstring of strings GACAATAA, ATAACAC, GTATA, TAATTGTA.
- Find the de Bruijn graph for k=2 (nodes will be pairs of nucleotides) and reads CCTGCC, GCCAAC
Sequence alignment
Main concepts in English and Slovak
The problem of local and global alignment of two sequences, dynamic programming algorithms, scoring matrix and its probabilistic meaning, statistical significance (E-value, P-value), heuristic search of local alignments (BLAST, minimizers), whole-genome and multiple alignments
Problém lokálneho a globálneho zarovnania dvoch sekvencií, jeho riešenie pomocou dynamického programovania, skórovacia matica a jej pravdepodobnostný význam, štatistická významnosť (E-value, P-value), heuristické hľadanie lokálnych zarovnaní (BLAST, minimizer-y), celogenómové a viacnásobné zarovnania
Simple questions for the exam
- Fill in the dynamic programing matrices for local and global alignment of sequences TACGT and CAGGATT, where the match has score +3, mismatch -1, gap -2. Reconstruct also the optimal alignments found by the dynamic programming algorithm.
- Compute the score of the alignment shown below using the scoring matrix shown below, gap opening penalty -5, gap extension penalty -2 for each additional base. Find a global alignment with a higher score for these two sequences and compute its score. (It is not necessary to find the optimal alignment; you can use any method to arrive at the answer.)
Alignment: Matrix:
ATAGTTTAA A C G T
A-GGG--AA A 2 -2 -1 -2
C -2 1 -2 -1
G -1 -2 1 -2
T -2 -1 -2 2
- Consider BLASTn algorithm starting from seeds of size w=3. How many seeds it finds while comparing sequences GATTACGGAT and CAGGATT? List all found seeds.
Gene finding
Main concepts in English and Slovak
Gene, exon, intron, mRNA, splicing and alternative splicing, genetic code, hidden Markov model (HMM), its states, transition and emission probabilities, use of HMMs in gene finding, Viterbi and forward algorithms
Gén, exón, intrón, mRNA, zostrih a alternatívny zostrih, kodón, genetický kód, skrytý Markovov model (HMM), jeho stavy, pravdepodobnosti prechodu a emisie, použitie HMM na hľadanie génov, Viterbiho a dopredný algoritmus
Simple questions for the exam
- What is the probability of generating sequence AGT using sequence of states 1,2,1 in the HMM below?
The HMM has three states 1, 2, 3. It always starts in state 1. Transition probabilities: From 1 to 1: 0.99 From 1 to 2: 0.01 From 2 to itself: 0.9 From 2 to 1: 0.05 From 2 to 3: 0.05 From 3 to itself: 0.99 From 3 to 2: 0.01 Emmision probabilities in state 1: A 0.25, C 0.25, G 0.25, T 0.25 Emmision probabilities in state 2: A 0.3, C 0.2, G 0.2, T 0.3 Emmision probabilities in state 3: A 0.2, C 0.4, G 0.3, T 0.1
Evolution and comparative genomics
Main concepts in English and Slovak
Phylogenetic tree (rooted and unrooted), maximum parsimony method, neighbor joining method, maximum likelihood method, Jukes-Cantor substitution model and more complex substitution matrices, Felsenstein algorithm, homolog, paralog, ortholog, detection of positive and negative selection, phylogenetic HMMs, likelihood ratio test
Fylogenetický strom (zakorenenený a nezakorenený), metóda maximálnej úspornosti (parsimony), metóda spájania susedov (neighbor joining), metóda maximálnej vierohodnosti (maximum likelihood), Jukes-Cantorov model substitúcií a zložitejšie substitučné matice, Felsensteinov algoritmus, homológ, paralóg, ortológ, detekcia pozitívneho a negatívneho výberu, fylogenetické HMM, test pomerom vierohodností (likelihood ratio test)
Simple questions for the exam
- Find the most parsimonious assignment of bases at the ancestral nodes in the tree below given a column of alignment TTAAA (in the order gollum, hobbit, human, elf, orc). You can derive your answer using any method (but it has to be the most parsimonious assignment).
Gollum ----|
|----|
Hobbit ----| |----|
| |
Human ---------| |
|---
Elf --------| |
|--------|
Orc --------|
- Find the most parsimonious tree for the alignment given below. What is its cost (i.e. how many mutations are necessary to explain these sequences)? You can derive your answer using any method (but it has to be the most parsimonious tree).
whitebird ACAACGTCT blackbird TCTGAATCA graybird TGTGAAAGA blubird ACTACGTCT greenbird TGTGAAAGA
- Consider the tree for gollum, hobbit etc. given above, where each branch has the same length t. Let us assume that for any two different bases x and y, the probability of base x mutating to base y over time y is 0.1, and thus the probability of base x remaining the same after time t is 0.7. Probability of each base in the root is 0.25. Compute the probability that the tree will have base A in all internal nodes and in leaves bases TTAAA (from top to bottom). Find an assignment of bases in the ancestral nodes with a higher probability and compute this probability (you do not need to find the best possible assignment).
- Consider the distance matrix given below. Which pair of nodes will be connected as first by the neighbor joining method and what will be the new distance matrix after joining these two nodes?
white black gray blue
whitebird 0 5 7 4
blackbird 5 0 8 5
graybird 7 8 0 5
bluebird 4 5 5 0
Gene expression, regulation, motifs
Main concepts in English and Slovak
Measuring gene expressing using microarray or RNA-seq, hierarchical clustering, classification, representation of sequence motifs (transcription factor binding sites) as a consensus, regular expression and PSSM, finding new motifs in sequences, consensus pattern problem, finding motifs using probability models, EM algorithm
Určovanie génovej expresie pomocou microarray alebo sekvenovaním RNA-seq, hierarchické zhlukovanie, klasifikácia, reprezentácia sekvenčných motívov (väzobné miesta transkripčných faktorov) ako konsenzus, regulárny výraz a PSSM, hľadanie nových motívov v sekvenciách, consensus pattern problem, hľadanie motívu pomocou pravdepodobnostných modelov, algoritmus EM
Simple questions for the exam
- After a series of expression measurements for 5 genes, we have computed distances between pairs of expression profiles and obtained the distance table shown below. Find the hierarchical clustering of these genes, where the distance between two clusters is computed as the minimum of the closest genes in these clusters (single linkage clustering). Show the order in which you were creating individual clusters.
A B C D E
gene A 0 0.6 0.1 0.3 0.7
gene B 0.6 0 0.5 0.5 0.4
gene C 0.1 0.5 0 0.6 0.6
gene D 0.3 0.5 0.6 0 0.8
gene E 0.7 0.4 0.6 0.8 0
- Consider a motif represented by the PSSM shown below. Compute the score of string GGAG. Which sequences of length 4 will have the smallest and highest score in this PSSM?
A -3 3 -2 -2 C -2 -2 1 -2 G 0 -2 -1 3 T 1 -1 1 -2
- We are running the EM algorithm for finding matrix W of base frequencies at individual motif positions. Motif length is 3 and we have 2 sequences of length 5 each, so there are 3 positions where the motif can start in each sequence. For each of these positions, we calculated the probability that the motif starts there based on the previous version of matrix W. What will be the new matrix W? (We do not use pseudocounts here, so for example base G will have probability 0 everywhere.)
Sequence 1: AACAT, probabilities of motif starting at positions 0,1,2: 0.4, 0.2, 0.4 Sequence 2: AAAAT, probabilities of motif starting at positions 0,1,2: 0.2, 0.2, 0.6
- Find all occurrences of regular expression TA[CG][AT]AT in sequence GACGATATAGTATGTACAATATGC.
Proteins
Main concepts in English and Slovak
Primary, secondary and tertiary structure of a protein, protein domains and families, family representation by a profile (PSSM) and a profile HMM, protein threading, gene ontology.
Primárna, sekundárna a terciálna štruktúra proteínov, proteínové domény a rodiny, reprezentovanie rodiny pravdepodobnostným profilom a profilovým HMM, protein threading, gene ontology.
Simple questions for the exam
- Construct a profile (PSSM) for the sequence alignment shown below, assuming that amino acid A comprises 60% of all sequences in a database, G 40% and we do not consider other amino acids. Use natural logarithm (ln) and pseudocount 1.
AAGA GAGA GAAA GGAG GGAA
RNA
Main concepts in English and Slovak
Secondary structure of RNA, pseudoknot and well-parenthesized structures, Nussinov algorithm, energy minimization, stochastic context-free grammars, covariance models of RNA families
Sekundárna štruktúra RNA, pseudouzol a dobre uzátvorkovaná štruktúra, Nussinovovej algoritmus, minimalizácia energie, stochastické bezkontextové gramatiky, kovariančné modely rodín RNA.
Simple questions for the exam
- Fill in the missing values in the matrix of dynamic programming for Nussinov algorithm which finds the RNA secondary structure without pseudoknots with the highest number of paired bases in RNA sequence GAACUAUCUGA (we allow only complementary bases A-U, C-G) and show the secondary structure found by the algorithm.
0 0 0 1 1 2 2 3 3 ? ?
0 0 0 1 1 2 2 3 3 ?
0 0 1 1 2 2 2 3 3
0 0 1 1 1 1 2 3
0 1 1 ? 1 2 3
0 1 1 1 2 2
0 0 0 1 2
0 0 1 1
0 0 1
0 0
0
- Consider RNA sequence of length 27, which has secondary structure pairs on positions (2,23), (3,22), (4,21), (5,13), (6,12), (8,16), (9,15), (10,14), (18,26), (19,25). What is the smallest number of pairs that needs to be removed from this list to get a structure without pseudoknots? Which pairs will be removed?
- Consider RNA sequence ACUGAGUCCAAGG, which has secondary structure pairs on positions (1,7), (2,6), (3,5), (8,13) a (9,12). (Positions are numbered started from 1.) (This RNA is shown as an example in the lecture on RNA structure.) Show a derivation of this sequence using a grammar show below so that paired bases are always generated in the same step of the derivation.
- Grammar 1: S->aSu|uSa|cSg|gSc|aS|cS|gS|uS|Sa|Sc|Sg|Su|SS|epsilon
- Grammar 2: S->aSu|uSa|cSg|gSc|TS|ST|SS|epsilon; T->aT|cT|gT|tT|epsilon
Population genetics
Main concepts in English and Slovak
Polymorphism, SNP, allele, homozygote, heterozygote, recombination, allele frequency as a Markov chain, random genetic drift, linkage disequilibrium, association mapping, LD block, subpopulation.
Polymorfizmus, SNP, alela, homozygot, heterozygot, rekombinácia, frekvencia polymorfizmu ako markovovský reťazec, náhodný genetický drift, väzbová nerovnováha (linkage disequilibrium), mapovanie asociácií, LD blok, subpopulácia.
Simple questions for the exam
- For pairs of SNPs from the tables show below determine, if we can statistically reject the hypothesis that they are in linkage equilibrium (LE) at the significance level p=0.05, i.e.
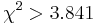 . For each pair compute their
. For each pair compute their 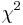 value.
value.
Q q Q q Q q
P 100 200 P 10 20 P 1 2
p 300 200 p 30 20 p 3 2
Additional important concepts
- Advanced dynamic programming algorithms (protein MS/MS, variants of sequence alignment, variants of Nussinov algorithm)
- Integer linear programming