CB05: Rozdiel medzi revíziami
Z MBI
| Riadok 13: | Riadok 13: | ||
Dalsi biologicky priklad HMM: topologia transmembranovych proteinov. | Dalsi biologicky priklad HMM: topologia transmembranovych proteinov. | ||
| − | == | + | ==E-hodnota (E-value) zarovnania== |
| − | + | * Priklady k tejto casti v prezentacii {{pdf|Cb-evalue}} | |
| + | * Mame dotaz dlzky m, databazu dlzky n, skore najlepsieho lokálneho zarovnania S | ||
| + | * E-value je ocakavany pocet zarovnani so skore aspon S ak dotaz aj databaza su nahodne | ||
| + | * Hrackarsky priklad: dotaz dlzky m=10, databaza dlzky n=300, S=6 | ||
| + | * Zoberme nas nahodny model s obsahom GC 50% | ||
| + | * Mame vrece s gulockami oznacenymi A,C,G,T, z kazdej 25% | ||
| + | * Vytiahneme gulicku, zapiseme si pismeno, hodime ju naspat, zamiesame a opakujeme s dalsim pismenom atd az kym nevygenerujeme m pismen pre dotaz a n pismen pre databazu | ||
| + | * Pre nase vygenerovane sekvencie spocitame, kolkokrat sa dotaz vyskytuje v databaze | ||
| + | * Cely experiment opakujeme vela krat a spocitame priemerny pocet vyskytov, co bude odhad E-value | ||
| + | |||
| + | Vypocet strednej hodnoty vzorcom namiesto simulacie (rychlejsie) | ||
| + | * zlozita matematicka teoria [https://www.ncbi.nlm.nih.gov/BLAST/tutorial/Altschul-1.html] | ||
| + | * E-value sa priblizne da odhadnut vzorcom: | ||
| + | :: <math>E = Kmn e^{-\lambda S}</math> | ||
| + | * n a m su dlzky porovnavanych sekvencii, S je skore, K a lambda su parametre, ktore zavisia od skorovacej schemy a od frekvencii vyskytu jednotlivych baz v nasom modeli nahodnej sekvencie. | ||
| + | * Napr blastn pre skorovaci system zhoda 1, nezhoda -1, medzera -2 používa lambda=0.800, K=0.0640 | ||
| + | * <math>E = 0.0640 nm 0.45^S</math>. | ||
| + | * Zdvojnásobenie dĺžky databázy alebo dĺžky dotazu zdvojnásobí E-value | ||
| + | * Zníženie skóre o 1 tiež zhruba zdvojnásobí E-value (delenie 0.45, t.j. nasobenie 2.2) | ||
| + | ** Cislo, ktorym nasobime, zalezi od lambda a teda od skorovacej schemy a frekvencii vyskytu baz | ||
Verzia zo dňa a času 07:24, 20. október 2021
Príklady stavových automatov pre HMM
Uvazujme HMM so specialnym zaciatocnym stavom b a koncovym stavom e, ktore nic negeneruju.
- Nakreslite HMM (stavovy diagram), ktory generuje sekvencie, ktore zacinaju niekolkymi cervenymi pismenami a potom obsahuju niekolko modrych
- Ako treba zmenit HMM, aby dovoloval ako "niekolko" aj nula?
- Ako treba zmenit HMM, aby pocet cervenych aj modrych bol vzdy parne cislo?
- Ako zmenit HMM, aby sa striedali cervene a modre kusy parnej dlzky?
V dalsich prikladoch uvazujeme aj to, ktore pismena su v ktorom stave povolene (pravdepodobnost emisie > 0) a ktore su zakazane
- cervena sekvencia dlzky dva, ktora zacina na A
- cervena sekvencia dlzky dva, ktora je hocico okrem AA
- toto sa da rozsirit na HMM, ktory reprezentuje ORF, teda nieco, co zacina start kodonom, potom niekolko beznych kodonov, ktore nie su stop kodonom a na koniec stop kodon
Dalsi biologicky priklad HMM: topologia transmembranovych proteinov.
E-hodnota (E-value) zarovnania
- Priklady k tejto casti v prezentacii pdf
- Mame dotaz dlzky m, databazu dlzky n, skore najlepsieho lokálneho zarovnania S
- E-value je ocakavany pocet zarovnani so skore aspon S ak dotaz aj databaza su nahodne
- Hrackarsky priklad: dotaz dlzky m=10, databaza dlzky n=300, S=6
- Zoberme nas nahodny model s obsahom GC 50%
- Mame vrece s gulockami oznacenymi A,C,G,T, z kazdej 25%
- Vytiahneme gulicku, zapiseme si pismeno, hodime ju naspat, zamiesame a opakujeme s dalsim pismenom atd az kym nevygenerujeme m pismen pre dotaz a n pismen pre databazu
- Pre nase vygenerovane sekvencie spocitame, kolkokrat sa dotaz vyskytuje v databaze
- Cely experiment opakujeme vela krat a spocitame priemerny pocet vyskytov, co bude odhad E-value
Vypocet strednej hodnoty vzorcom namiesto simulacie (rychlejsie)
- zlozita matematicka teoria [1]
- E-value sa priblizne da odhadnut vzorcom:
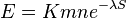
- n a m su dlzky porovnavanych sekvencii, S je skore, K a lambda su parametre, ktore zavisia od skorovacej schemy a od frekvencii vyskytu jednotlivych baz v nasom modeli nahodnej sekvencie.
- Napr blastn pre skorovaci system zhoda 1, nezhoda -1, medzera -2 používa lambda=0.800, K=0.0640
-
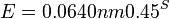 .
.
- Zdvojnásobenie dĺžky databázy alebo dĺžky dotazu zdvojnásobí E-value
- Zníženie skóre o 1 tiež zhruba zdvojnásobí E-value (delenie 0.45, t.j. nasobenie 2.2)
- Cislo, ktorym nasobime, zalezi od lambda a teda od skorovacej schemy a frekvencii vyskytu baz